
. Introduction
Ecologists, since more than a century, have long been interested in ecotones (Clements, 1905; Odum, 1953). Ecotones, representing transition zones between different adjacent ecological regions, play a fundamental role in generating species biodiversity and evolutionary novelty, thus deserving particular attention in monitoring, managment and conservation programs (Smith et al., 1997; Farina, 2008; Kark, 2013).
Riparian zones, where terrestrial and freshwater ecosystems meet, are a classic example of ecotone (Naiman and Decamps, 1997). These interfaces are among the most diverse and dynamic biophysical habitats on earth, and are particularly sensitive to environmental changes (Naiman and Decamps, 1997).
In riparian areas, vegetation contributes to carbon storage, serves as wildlife habitat and ecological corridor, stabilizes streambank and provides shade, food and organic matter for stream and its biota (Salemi et al., 2012). Riparian forests are therefore recognized for their ecological importance, and the understanding of their growth dynamics and its relation with environmental variability is mandatory in the context of a climatic unstable scenario (IPCC, 2007).
Along arid and semi-arid areas worldwide, different species belonging to the Prosopis genus (Fabaceae) play a key role in ecosystem structuring (Villagra et al., 2010). This is the case of P. flexuosa DC (algarrobo dulce), a facultative phreatophyte tree species dominating the Central Monte Desert woodlands (central-west Argentina). In this region, the species grows in territories characterized by a permanent access to water reservoirs, thus where groundwater is relatively shallow (6–15 m depth), or along riverbanks (Morello, 1958), where the P. flexuosa riparian forests contribute to sustain local human development and activities (Goirán et al., 2012).
Dendrochronology is a useful tool in analyzing tree and shrub growth dynamics (Fritts, 1976). Since the ring width represents a proxy record of different abiotic and biotic forcings influencing radial growth, dendrochronological methods have been successfully applied in analyzing the influence of hydrological factors upon riparian species stem growth (e.g. Heuzé et al., 2009; Rodríguez-González et al., 2014).
A number of studies have explored the radial growth dynamics of P. flexuosa woodlands in the Central Monte Desert, analyzing the relation between tree-ring growth and climatic factors as well as the role of environmental heterogeneity upon the ring development (e.g. Giantomasi et al., 2012, 2013; Piraino et al., 2015). Nevertheless, a comprehensive understanding of the species sensitivity to hydrological changes in riparian settings is still lacking (but see Piraino et al., 2015, for a first description of riparian P. flexuosa growth and its relation with precipitation variability). Furthermore, to our knowledge, while several researches examined the relation between hydrology and Prosopis forests stand structure and biomass (see for instance Stromberg et al., 1993), no study has been performed thus far regarding the dendrohydrology of any Prosopis species. To fill this gap of knowledge, we applied dendrochronological techniques to several P. flexuosa trees distributed in a riparian area located in the Central Monte Desert district, differing in their spatial position relative to the riverbank. We hypothesize that i) the species radial growth is mainly determined by hydrological rather than atmospheric climatic factors; ii) growth-hydrological relations are modulated by tree location in the riparian area. Due to the widespread distribution of Prosopis spp. in drylands, with this research we aimed to contribute to the understanding of arid riparian forest growth dynamics, and furthermore stressing the potential utility of the Prosopis genus in dendrohydrological studies.
. Material and Methods
Sites description
The study area is located in the Central Monte Desert district belonging to the Mendoza Province, central-west Argentina (Fig. 1). Climate is arid to semi-arid, characterized by a mean annual precipitation of 155 mm, and by large seasonal and daily temperature variability (Morello, 1958; Abraham et al., 2009) (Fig. 1).
Fig. 1
Geographical location of the study area (a), spatial position of the sampled plots (b), and Ombrothermic diagram drawn according to the methods of Bagnouls and Gaussen (1953) (c). Precipitation and temperature data belongs to the Encón climatic station (32° 15’ S, 67° 47’ W), covering the 1971–1987 period. Source for the satellite images was Google Earth http://earth.google.es.
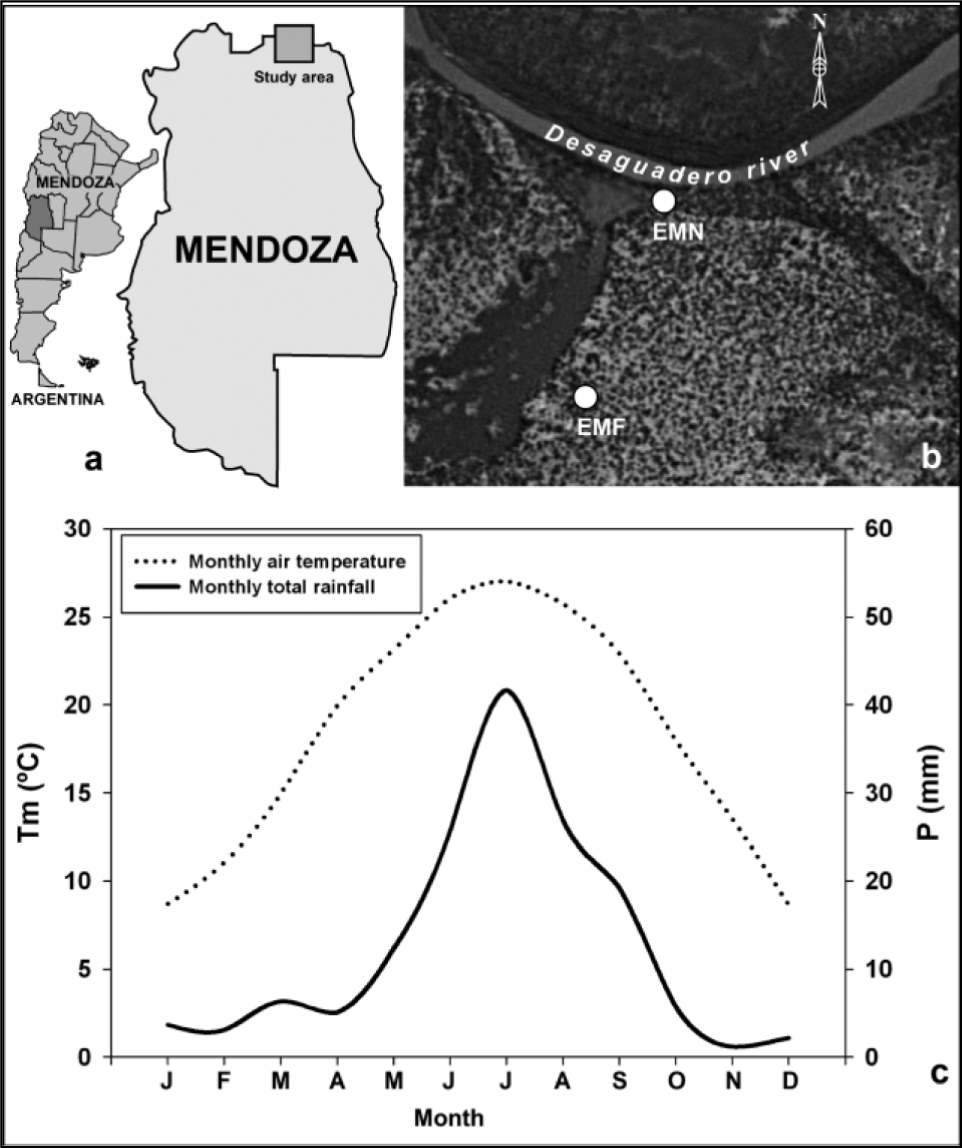
We selected one site (El Mateo) of P. flexuosa forest growing near the Desaguadero river (32°14′31.4″S; 67°41′21.8″W; Fig. 1). At the analyzed forest stand, vegetation is composed of P. flexuosa with shrub and low arboreal associates such as Larrea divaricata Cav., Geoffroea decorticans (Hook. & Arn.), Capparis atamisquea Kuntze, and Bulnesia retama (Hook.) Griseb. Sampled site presents evidence of relatively recent human pressure (stumps), as most of the P. flexuosa woodlands distributed in the Central Monte Desert (Villagra et al., 2009).
During the austral winter of 2011, 2 rectangular plots of 1000 m2 (50 × 20 m), differing in hydrological settings (EMN: El Mateo near; EMF: El Mateo far; Fig. 1), were established, with the longest plot side parallel to the streambank (Table 1). Within the 2 plots, dendrochrono-logical sampling was conducted on all trees regardless of their one- or multi-stemmed form. One to two cores per tree were extracted at about 50 cm above the ground with a gas-powered drill (TED_262R, Tanaka Kogyo Co. Ltd, Chiba, Japan) from a total of 62 trees.
Table 1.
Geographical and hydrological settings of the sampled plots. EMN — El Mateo near; EMF — El Mateo far. DRB — plot distance from the riverbank. GWD — groundwater depth.
| Plot | Altitude (m) | DRB (m) | GWD (m) | Latitude (°S) | Longitude (°W) |
|---|---|---|---|---|---|
| EMN | 514 | 15 | 2.9 | 32°14′18.3″ | 67°41′14.7″ |
| EMF | 513 | 400 | 3.6 | 32°14′30.7″ | 67°41′21.8″ |
Standard dendrochronological procedures followed Stokes and Smiley (1968): wood samples were glued on wooden supports and polished with progressively finer sandpaper (60–1200 grit), then tree-ring were identified and dated, and widths were measured from pith to bark to the nearest 0.01mm resolution through the facilities of the IPWin4 Image Analysis software (v4.5, Media Cybernetics, USA). The obtained tree-ring series were crossdated and their data quality validated by statistical controls (COFECHA program; Holmes, 1983).
The age-related trend and the influence of exogenous disturbance recorded in raw ring width measurements were removed by standardization procedures through the program ARSTAN40c (Cook and Krusic, 2006). We chose a double de-trending method (Cook, 1985): first, a negative exponential function was fit to each raw tree-ring series, and then we applied a 32-year cubic spline function with 50% cut-off. To reduce the autocorrelation present in ring-width series, autoregressive model of order determined by the Akaike Information Criterion (AIC; Sakamoto et al., 1986) was applied. Tree-ring indices were computed by dividing the measured ring widths by the expected value. Finally, individual standardized series were averaged to produce a mean chronology for each site. We selected the “Residual” version standardized chronology obtained from the ARSTAN40c program, which avoids any possible overestimation of the climate-growth relation (Villalba and Veblen, 1997).
The internal quality of each chronology was examined according to the following statistical indexes: MS (Mean Sensitivity), a measurement of the relative year-to-year change in tree-ring width; EPS (Expressed Population Signal), an estimation of how well a finite-sample chronology can represent the theoretical population chronology based on an infinite number of trees; and RBAR, a measure of the common variance between the single series in a chronology (Fritts, 1976; Wigley et al., 1984). The two latter indexes were calculated for a 15-year win-dow with a 14-year overlap. EPS, following a previous study performed for P. flexuosa woodlands distributed in the Central Monte Desert (Piraino et al., 2015), was considered satisfactory when surpassing a threshold of 0.75 instead of the original 0.85 value proposed by Wigley et al. (1984).
Statistical analysis
Environmental-growth relations were calculated through a combination of different dendroecological and statistical methods, considering streamflow and precipitation as the abiotic drivers of the species stem growth.
First, since rapid environmental changes are supposed to cause the formation of extremely narrow or wide tree-ring as response to particularly adverse or favorable conditions, raw annual growth series were examined through pointer year analysis (Schweingruber et al., 1990). Pointer years were calculated following the “normalization in a moving window” method, by selecting a 5-years-long window (Cropper, 1979). A given year was considered as a pointer one when at least 67% of the analyzed trees exhibited a change in the radial growth greater than 50%. Calculations were performed through the WEISER software (Gonzalez, 2001). Once established, pointer years were compared to annual streamflow and rainfall averages, respectively defined according to hydrological and biological criteria. Mean annual streamflow, coincident with previous year July-current year June period, was computed following Vich et al. (2014). On the other hand, annual precipitation average corresponded to previous year April-current year March period, representing the time window between the end of preceding and current growing seasons (Giantomasi et al., 2012).
Then, to assess the most important hydrological factors influencing radial growth, correlation functions (Blasing et al., 1984) were performed matching residual ring growth against monthly streamflow and precipitation amount. In order to directly compare the influence of both hydrological variables upon the ring development, we focused on the common time period from preceding year April to current year March, when cambium activity ends (Giantomasi et al., 2012). Correlations were computed with the software DENDROCLIM2002 and tests for significance were determined through the bootstrap method (Guiot, 1991; Biondi and Waikul, 2004). Possible differences in radial growth-environmental variability relation as function of tree spatial distribution were analyzed by the means of ANOVA computed on the r/s coefficients (r being the Pearson correlation moment, and s its standard deviation), focusing only on the current growing season period (October to March; Giantomasi et al., 2012).
Finally, regression analyses were computed to model the relation between radial growth and environmental variables. For model selection, we considered an information-theoretic approach, using Akaike Information Criterion corrected for small samples (AICc; Burnham and Anderson, 2002) to compare a suite of competing models. The best model was selected using the lowest AICc value from a set of candidates. AICc was calculated through the “AICcmodavg” package present in the R software (R Core Development Team 2011; Mazerolle and Mazerolle, 2015).
In all the abovementioned analyses, calculations were performed for the common time period 1991–2008 (n = 18), selected due to the short extension of stream-flow data (available for 1991–2012), and to the last year (2008; data not shown) of both EMN and EMF tree-ring chronologies showing an EPS value higher than the 0.75 threshold established in this research. Although precipitation data covers a larger period (1950–2011), through the established common time window we obtained a clearer picture of the dendrohydrological potential of the studied species. Streamflow data, belonging to the Encón gauge station (32°13′30.10″ S; 67°48′24.00″ W), located at approximately 12 Km from the analyzed forest stand, were downloaded from the webpage of the “Subsecretatia de Recursos Hídricos” (available at http://www.hidricosargentina.gov.ar/acceso_bd.php). Precipitation values were obtained from the San Juan Airport gauge station (31° 34’ S; 68° 25’ W), placed at 100 Km from the study area and which have been successfully used in a previous dendroclimatological analysis of P. flexuosa (Piraino et al., 2015).
. Results
Characteristics of the tree-ring chronologies
As reported in previous researches performed in the Central Monte Desert, the P. flexuosa wood is particularly heavy and hard, and difficult to cut (Villalba, 1985; Iglesias, 2010). Annual rings are semi-ring-porous and present small to large-sized vessels with simple perforation plates (Villalba, 1985). Tree-ring structure is more evident in the radial section close to the pith, since closer to the bark rings become thinner and more difficult to recognize (Giantomasi et al., 2009). Nevertheless, the transition from early- to late-wood, characterized by a decrease in average pore diameter, along with the presence of abundant axial parenchyma, allowed a reliable determination of the annual species radial growth (Fig. 2).
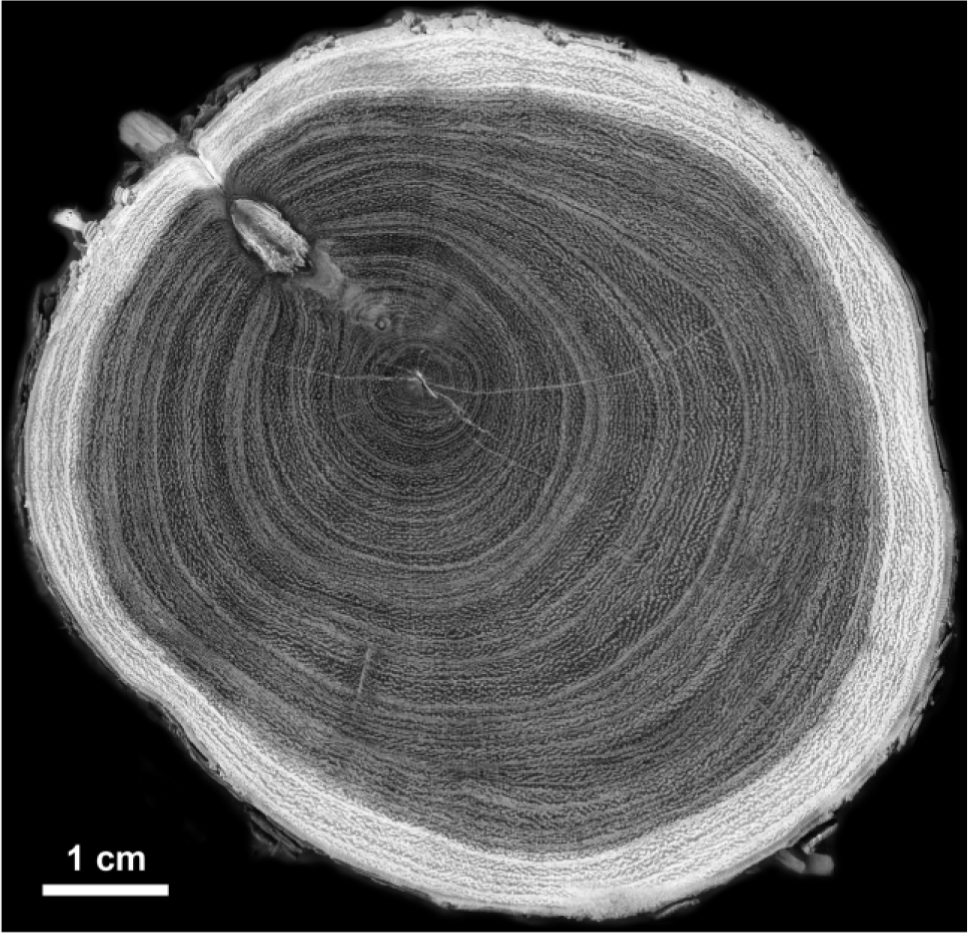
38 samples from 32 trees contributed to the construction of the two tree-ring chronologies, spanning from 47 to 51 years (Table 2). During the chronologies development, two trees belonging to the EMF plot were excluded, in order to obtain radial growth data from individuals of similar age and therefore reduce the ‘noise’ caused by the variation of other factors than spatial position in the riparian zone. Mean annual growth rate, mean correlation (MC), mean sensitivity (MS), as well as mean EPS and RBAR values present similar results among the developed chronologies, being slightly higher for the EMN plot (Table 2).
Table 2.
Characteristics of the tree-ring chronologies. N — Number of sampled trees per plot (in parenthesis: total number of dendrochrono-logical samples); Period — time range of the sampled cores; TRW — mean ring-width value; MC — mean correlation between series at each plot; MS — mean sensitivity; EPS — mean Expressed Population Signal, RBAR — average correlation between all series.
| Plot | N | Period | TRW (mm) | MC | MS | EPS | RBAR |
|---|---|---|---|---|---|---|---|
| EMN | 19(21) | 1964-2010 | 1.56 | 0.518 | 0.470 | 0.74 | 0.16 |
| EMF | 13(17) | 1960-2010 | 1.52 | 0.505 | 0.419 | 0.58 | 0.12 |
Streamflow and precipitation influences on tree radial growth
Pointer years analysis detected respectively three (years 1996, 2000 and 2007) and two (1996 and 2002) characteristics narrow tree-rings for the EMN and the EMF chronologies (Fig. 3). During 1996, at both EMN and EMF plots the totality of individual chronologies showed extremely narrow rings. Regarding the two remaining pointer years emerged at the EMN, 69% (2000) and 73% (2007) of the sampled trees experienced a strong decrease in their radial growth, while at the EMF plot, pointer year during 2002 where registered by 69% of the sampled individuals.
Fig. 3
Raw ring-width series of the analyzed trees showing pointer years (vertical lines) at the EMN and EMF plots for the common period 1991–2008.
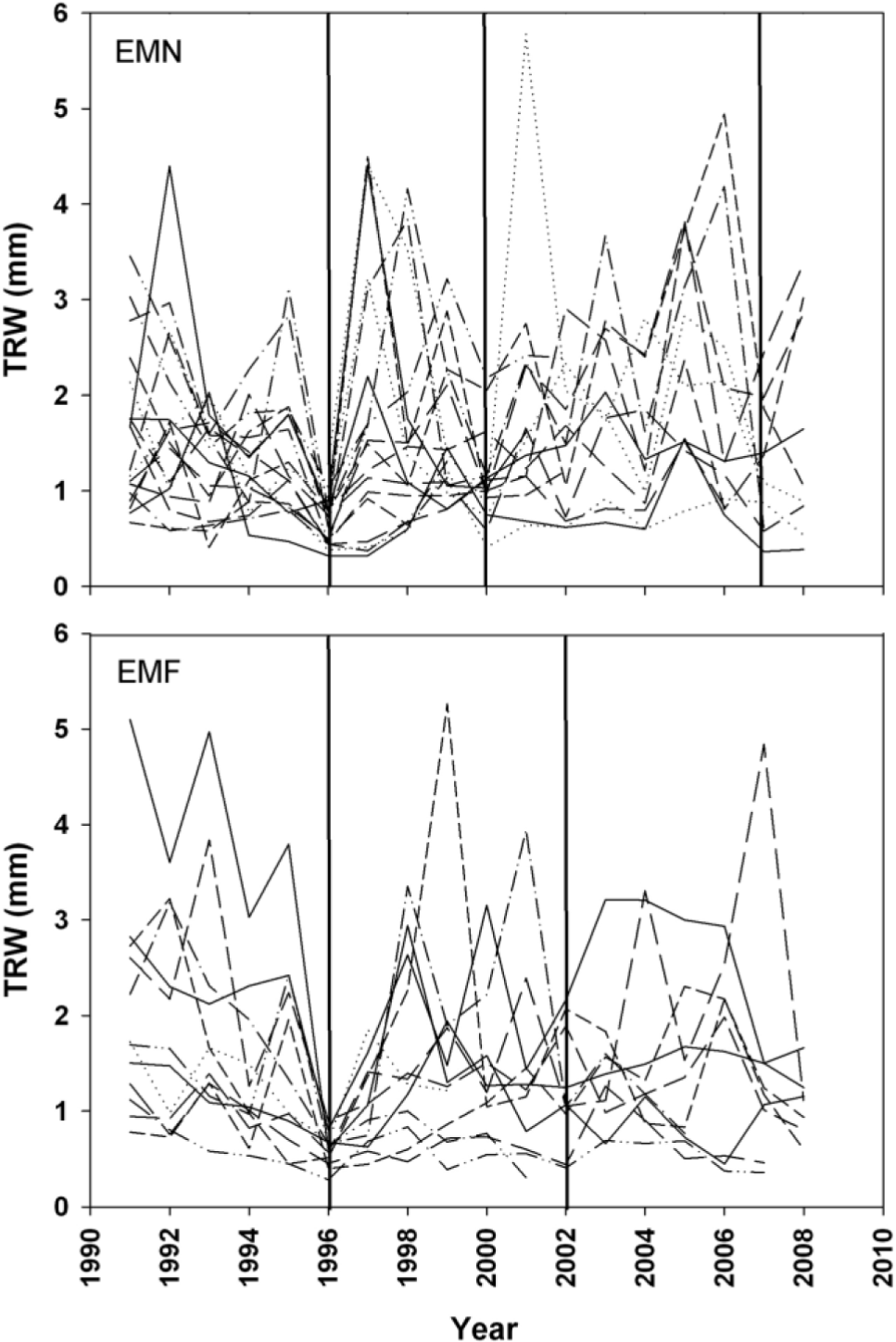
Narrow rings emerged for both the EMN and the EMF chronologies during 1996 corresponded to a year of zero-streamflow value. Concerning the EMN plot, annual streamflow values during the two remaining pointer years were lower than mean (years 2000 and 2007, respectively 34% and 89% of annual streamflow average), while annual precipitation was higher than historical mean (respectively 112% and 199%). Finally, the negative pointer year emerged during 2002 for the EMF chronology corresponded to a year characterized by lower than mean annual precipitation (94%) and higher than mean annual streamflow amounts (304%).
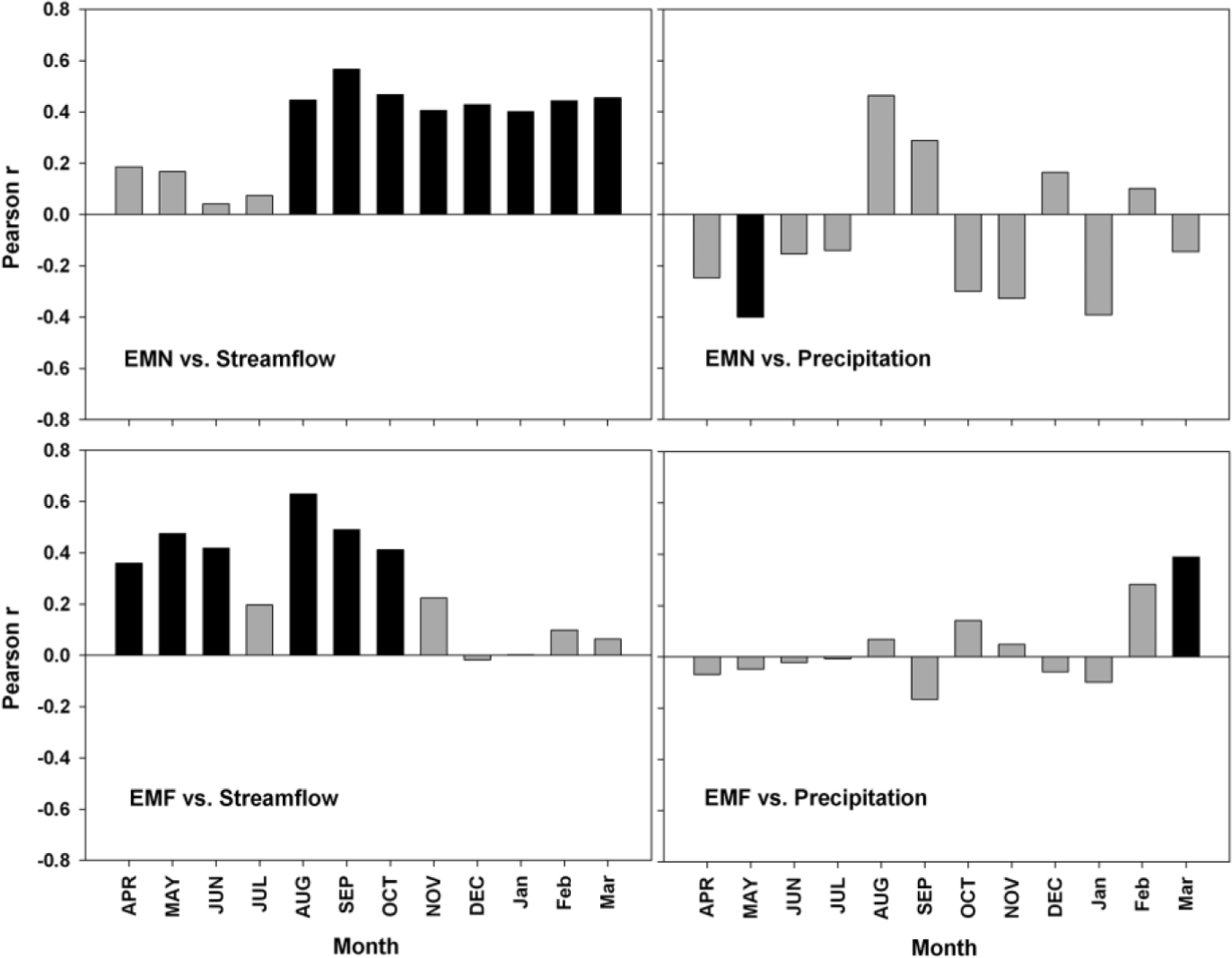
Correlation function analysis performed between standardized chronologies and monthly environmental data revealed different dendrohydrological signals among the selected P. flexuosa populations. EMN radial growth is favored by above-mean spring-summer streamflow values (September to March), while abundant precipitation during May negatively influenced the ring development (Fig. 4). Regarding the EMF chronology, ring growth is positively associated to high streamflow values during autumn and winter months (Fig. 4). Furthermore, at this plot radial growth is benefited by higher than mean streamflow values during spring period (September-October) and by late-summer (March) abundant precipitation (Fig. 4).
Fig. 4
Correlation functions between each EMN and EMF residual chronologies and monthly total streamflow and precipitation data for the common period 1991–2008. Black bars indicate significance at p<0.05.
Variability in the hydrological settings among the analyzed plots modulated the dendrohydrological signals regarding streamflow influence on the ring growth (oneway ANOVA, F = 17.08; p = 0.0002; data not shown), while no significant differences emerged concerning the precipitation-radial growth relation (one-way ANOVA, F = 4.68; p = 0.0557; data not shown).
Model selection using different combination of dependent and explanatory variables showed that the set of models for residual growth values depending on stream-flow and precipitation were stronger than the null model (no predictors; data not shown). EMN ring growth is best explained by a quadratic regression with mean stream-flow values of spring-summer months (September, December, January, February and March) as explanatory variable (Table 3). On the other hand, the EMF radial growth is best explained by a linear model with August streamflow and March precipitation as explanatory variables (being the last variable marginally statistical significant at the p<0.05 level; Table 3).
Table 3.
Model parameters (explanatory variables), significance (p), slope and adjusted coefficient of determination (R2adj.) for the “optimal model” of each dependent variable. StrFI — Streamflow; P — Precipitation. SDJFM referred to the mean streamflow values of September, December, January, February and March months.
| Plot | Constant | Parameter | p | Slope | Parameter | Slope | p | R2adj. |
|---|---|---|---|---|---|---|---|---|
| EMN | 0.9867 | StrFI(SDJFM) | 0.0222 | >0 | StrFI(SDJFM)2 | <0 | 0.0464 | 0.3516 |
| EMF | 0.791567 | StrFI(August) | 0.00731 | >0 | P(March) | >0 | 0.06887 | 0.416 |
. Discussion
At the two examined plots, raw tree-ring statistics (namely MC, MS, EPS and RBAR) showed slightly higher values for the EMN (nearest trees) than the EMF (farthest trees) chronology. This can be consequence of possible differences in hydrological settings among the P. flexuosa stands. In these sense, the results emerged in a previous research (Piraino et al., 2015) suggest that the proximity to a source of stable water supply, as for example an active stream, can influence the common ring growth signal, due to the minimization of the individualistic response to water-table fluctuations in a facultative phreatophytic species as P. flexuosa (Morello, 1958; Cook, 1987; Jobbágy et al., 2011).
Pointer years seemed to be mainly associated to streamflow rather than precipitation changes, although some differences emerged among the selected plots. Indeed, while the presence of narrow rings at both chronologies, experienced by 100% of the analyzed trees during 1996, corresponded to a year of zero-streamflow values, the remaining negative pointer years seemed to reflect a strongest dependence of nearest trees radial growth to streamflow extremes, while for farthest trees apparently the combination of both hydrological resources drove the occurrence of narrow ring widths.
Correlation function analysis results showed differences in the dendrohydrological signal among the sampled stands. At the EMN plot, radial growth is enhanced by above-average spring-summer streamflow values, and no statistically significant result emerged regarding growing season precipitation influence on the stem growth. On the other hand, at the EMF the ring development is positively related to winter and spring river discharge and late-summer precipitation amount.
From a physiological point of view, these results are understandable considering the different phases of the species cambium activity, and possible diverse strategies in sub-surface (streamflow) and meteoric (precipitation) water use, the latter probably reflecting the spatial distribution of the examined trees (Naiman and Decamps, 1997). In the Central Monte Desert, the P. flexuosa treering development takes place from October to March (Giantomasi et al., 2012). Thus, at the EMN plot, radial growth seemed to be tightly coupled to streamflow changes along the whole growing season. On the other hand, at the EMF plot, winter streamflow variability probably contributed to the recharge of water-table, thus favoring tree growth in the following growing season. Furthermore, for far trees, the activation of cambium possibly relied on above-average streamflow during the month of October, while the presence of a second pulse of radial growth, which takes place during February-March as evidenced by Giantomasi et al. (2012), is likely function of late-summer rainfall amount. In this sense, farthest trees are apparently more flexible in using different water sources than nearest trees, in agreement with an isotopic analysis performed for other riparian Prosopis species (P. vellutina) distributed in semi-arid regions of USA (Snyder and Williams, 2000).
The regression analysis further contributed to understand how the examined riparian P. flexuosa stand responds to streamflow changes, particularly for trees distributed near the riverbank. The quadratic model emerged for the EMN radial growth suggested that although a positive relation among the ring development and spring-summer streamflow exists (as found in correlation function analysis), an excessively abundant river discharge (e.g. flooding) can be detrimental for the species stem growth. This result evidenced that the EMN P. flexuosa radial growth can be inhibited by stress under conditions that are too wet or too dry. Thus, following the subsidy-stress theory, high river discharge can have both a positive and a negative effect upon tree growth, due to high water and nutrient availability (subsidy), or through long-term soil saturation and root anoxia (stress) (Odum et al., 1979; Rodríguez-González et al., 2010). This result is sound with studies analyzing the response of riparian temperate species such as Hedera helix L. and Taxodium distichum (L.) Rich. to hydrological changes (Heuzé et al., 2009; Palta et al., 2012).
Several works highlighted a heterogeneous response of riparian species radial growth to hydrological factors as function of hydrological settings variability (e.g. distance from the river bank, elevation, groundwater depth). Tardif and Bergeron (1993) analyzed the relation of Fraxinus nigra Marshall radial growth in a Canadian boreal floodplain with climatic and hydrological fluctuations. Trees distributed in sites exposed to flooding responded much more to the negative impact of spring precipitation, while differences in elevation influenced the species growth response to summer precipitation. Rodríguez-González et al. (2014) demonstrated that the sensitivity to hydrological changes of Alnus glutinosa (L.) Gaertn. in the eastern Iberian peninsula is function of differences in elevation and distance from the riverbank. Helama et al. (2013) examined the dendroclimatic signals of riparian and upland pines forest interior in Finland, and showed that the former reacted negatively to increased rainfall in June, whereas the latter showed a positive response to the same climatic forcing. Dudek et al. (1998) founded that differences in elevation modulate the response of Populus deltoids W. Bartram ex Humphry Marshall and Juglans nigra L. to stream-flow and climate. Regarding arid riparian forests, the strength in the flow-growth relationship of riparian Populus trichocarpa (Torr. & Gray) stands distributed in Sierra Nevada (USA) varied with tree position in the floodplain (Stromberg and Patten, 1996), while the eastern white pine (Pinus strobus L.) growth response to environmental variability is modulated by differences in hydrological settings among floodway and terrace sites (Chhin et al., 2013).
. Conclusions
In this research, we explored for the first time the role of stream-flow as a potential influencing factor on the radial growth variability of riparian Prosopis woodland. The presented results evidenced a strong dependence of the P. flexuosa radial growth to stream-flow changes, although some differences in the dendrohydrological signal as function of tree spatial distribution emerged. This information is particularly relevant considering the projected decrease in winter precipitation amount for the studied area (Boulanger et al., 2006), which will probably be detrimental for the forest growth dynamics. Finally, the relative high agreement between year-to-year growth variability and river discharge values suggested that P. flexuosa (and potentially other species belonging to the Prosopis genus) is a good candidate in dendrohydrological studies.